| The Perth Group
The HIV-AIDS debate | 
|
| The Perth Group
The HIV-AIDS debate |
|
 Home What the Perth Group has argued
Papadopulos redox theory of cellular function papers Papers and letters published in scientific journals Monograph on mother-to-child transmission
Papers published in Continuum magazine
Papers published in the popular press
Papers/letters rejected by the scientific press
Presentations
Interviews
Selected email correspondence
A virus like no other
Oxidation, Montagnier and the Perth Group Home What the Perth Group has argued
Papadopulos redox theory of cellular function papers Papers and letters published in scientific journals Monograph on mother-to-child transmission
Papers published in Continuum magazine
Papers published in the popular press
Papers/letters rejected by the scientific press
Presentations
Interviews
Selected email correspondence
A virus like no other
Oxidation, Montagnier and the Perth Group
 Montagnier Nobel Prize 2008
The Parenzee Case
The House of Numbers
Latest files
National Libary of Australia Intervew 1993
Others
Africa/South Africa
Questions and answers
Response to the NIH "Evidence" that HIV causes AIDS
Translations of the Perth Group papers
BMJ Online Debate
Contact Us
About the Perth Group
Perth Group at Virusmyth
The Perth Group on YouTube Montagnier Nobel Prize 2008
The Parenzee Case
The House of Numbers
Latest files
National Libary of Australia Intervew 1993
Others
Africa/South Africa
Questions and answers
Response to the NIH "Evidence" that HIV causes AIDS
Translations of the Perth Group papers
BMJ Online Debate
Contact Us
About the Perth Group
Perth Group at Virusmyth
The Perth Group on YouTube
|
Download the Western blot graphic HERE A CRITICAL ANALYSIS HIV SEROLOGY DOES NOT AFFIRM A RETROVIRAL INFECTION Rejected by Emergency Medicine [Australia] June 2002 Eleni Papadopulos-Eleopulos Biophysicist, Department of Medical Physics, Royal Perth Hospital, Perth, Western Australia Valendar F. Turner Consultant Emergency Physician, Department of Emergency Medicine, Royal Perth Hospital, Perth, Western Australia John M Papadimitriou Professor of Pathology, University of Western Australia, Perth, Western Australia Helman Alfonso Department of Research, Universidad Metropolitana Barranquilla, Colombia Barry A. P. Page Physicist, Department of Medical Physics, Royal Perth Hospital, Perth, Western Australia David Causer Physicist, Department of Medical Physics, Royal Perth Hospital, Perth, Western Australia Sam Mhlongo Head & Chief Family Practitioner, Family Medicine & Primary Health Care, Medical University of South Africa, Johannesburg, South Africa Todd Miller Research Assistant Professor, Department of Medicine, Division of Cardiology, University of Miami School of Medicine, Florida, United States of America Christian Fiala Gynaecologist, Department of Obstetrics and Gynaecology, General Public Hospital, Korneuburg, Austria Anthony Brink Advocate of the High Court of South Africa, Capetown, South Africa Neville Hodgkinson Science Writer, Oxford, England Manu Kothari Professor of Anatomy, Seth Gordhandas Sunderdas Medical College, King Edward Memorial Hospital, Mumbai, India
=============================================================== Man prefers to believe what he prefers to be true.Francis Bacon (1561-1626) AbstractClinicians and especially emergency physicians serve a principal role in the recognition and management of patients with or at risk of developing the acquired immune deficiency syndrome (AIDS). The scientific basis for this responsibility rests on the detection of antibodies which react with particular proteins claimed to be those of a unique, exogenous retrovirus, HIV. It remains to be irrefutably proven that such antibodies or proteins bear any relationship to infection with a retrovirus.
IntroductionIn 1890 von Behring and Kitazato discovered that laboratory animals exposed to bacteria developed substances in their sera capable of restoring the health of other infected animals.1 It was postulated that bacterial toxins �under certain conditions, will lead the organism to produce substances which render them harmless and which prevent the development of the bacteria�.2 As a result, �antitoxins� were taken from laboratory to bedside and proved successful for the treatment of children dying from diphtheria. Subsequently von Behring was awarded the first Nobel Prize in Physiology and Medicine for his work on serum therapy. The substances believed responsible for the therapeutic benefit became known as Antik�rper (antibodies) and nowadays agents inducing their synthesis are encompassed under the generic title antigens (ANTIbody GENerating). Although the precise relationship between antigens, antibodies and immunity remains elusive, antigen/antibody interaction has found widespread acceptance as an indirect means of determining infection with a particular microbe. Indeed, data of this type are quintessential to the construction of the HIV theory of AIDS.
Problems with the interpretation of antigen/antibody reactivityvon Behring and Kitzato�s interpretation of the existence of Antik�rper directed against bacterial toxins was contingent upon their prior knowledge that certain laboratory animals had been exposed to bacterial antigens. In clinical practice the order of data is inverted, that is, from the detection of antibodies clinicians seek proof of exposure to an antigen. Unfortunately, Nature has conspired against the clinician by not affording the straightforward interpretation available to von Behring and Kitzato. Although antigen �X� may be defined beforehand by other means and with the utmost precision, from an antibody test all a scientist may conclude is the presence or absence of a reaction. For several reasons, from this data alone it is incorrect to deduce that the antibody arises because of, or is directed against, antigen �X�. Additional steps must be undertaken before such a conclusion is vindicated. The explanation for this relates to three factors:
Identical antigens from dissimilar sources Similar antibodies, even if specifically directed against a given antigen, may arise if the antigen is present in dissimilar sources. For example, cardiolipin is a phospholipid found in cell nuclei, inner mitochondrial membranes and the plasma membranes of bacteria. Antibodies which react with this antigen appear following infection with T. pallidum and are demonstrated by the classical Wasserman, Kahn and VDRL reactions. However, similar reactivity occurs in numerous other conditions constituting the well known causes of false-positive syphilis serology. A similar mechanism underlies the Weil-Felix reactions for Rickettsial organisms using Proteus OX-19 and OX-2 strains.
Cross-reactivity According to Nossal antibodies �are highly specific molecules. For each antigen there is a corresponding different antibody�. Yet also according to Nossal, �this is not the case�an antibody molecule made following the injection of one antigen frequently can combine also with a second antigen�in other words, the antibody cross-reacts with the second antigen�It is simply because the fit between antigen and antibody need not be 100 per cent perfect that the body can turn out antibodies to virtually everything�On its bountiful key-ring the body will find at least one key which fits sufficiently well to open any door�.3 Cross-reactivity is a property of all antibody molecules4-12 including monoclonal antibodies6, 10, 13 and there are instances where "cross-reactive antibodies may have higher affinity with antigens other than the inducing antigen�.12 All that one needs to be convinced that all "antibodies are polyspecific, that is, they are able to react with various dissimilar antigens such as: proteins, nucleic acids and haptens" and "they are able to react with more than to self or non-self antigens, often without any apparent antigenic similarities", is to read the scientific publications of the researchers, such as Stratis Avrameas, from the Pasteur Institute.6 A germane example of cross-reactivity is a study of acute measles infection in non-HIV infected children and young adults reported from the USA and Peru. Here 43/75 (62%) of patients developed antibodies which reacted with one or more of the proteins present in the HIV test kits.14 (In 1995 in Africa there were 551,000 childhood deaths from measles15 and awareness of this problem may explain why, in one African study of HIV infection, �Children hospitalized with measles were excluded�16). Failure to heed cross-reactivities is akin to pronouncing an unknown substance sodium chloride merely because its admixture in solution with silver nitrate results in a white precipitate. If this were the case analytical chemistry could be mastered in a matter of days. Few if any substances react monogamously and this property of matter pervades not only chemistry but also serology.
Non-specific induction Antibody production may also be induced non-specifically,17-19 that is, antibodies directed against a defined antigen arise following exposure to an unrelated antigen. The classical example is that of Epstein-Barr virus infection which results in polyclonal B-cell stimulation and a large repertoire of antibodies including heterophile antibodies directed against sheep and horse erythrocytes.19 This example further illustrates the central problem of serological practice. A scientist may predict that an experiment where humans are injected with sheep or horse erythrocytes will result in the production of antibodies directed against these antigens. However, reactivity with these antigens, revealed for example in a patient by a Monospot� test, is not interpreted as proof of infection with animal red blood cells, or that infectious mononucleosis is caused by these agents. Rather it exemplifies the absolute necessity of proving the specificity of antigen/antibody reactions for whatever antigen is sought before they are introduced into routine clinical practice.
The HIV antibody tests There are two "HIV" antibody tests in common use, the ELISA and Western blot (WB). In the ELISA a colour change occurs when a mixture of the proteins attributed to HIV reacts with antibodies in serum from a patient. The same principal applies to all generation ELISAs including the newer agglutination, immunofiltration (flow through tests) immunochromatographic (lateral flow tests) and dipstick tests. The significant difference between the �screening� or �initial� ELISA and the �supplemental� or �confirmatory� Western blot is that in the latter the test proteins are electrophoretically separated along the length of a nitrocellulose strip. This enables reactions between antibodies and individual proteins to be visualized as a series of bands. These are referred to with a small �p� (for protein) followed by a number designating the protein�s molecular weight in thousands, for example, p24. In the majority of countries the diagnosis of HIV infection consists of performing an initial ELISA which, if reactive, is repeated. Repeatedly reactive ELISAs are �confirmed� by a positive Western blot which is regarded as synonymous with HIV infection. In the United States �Reactive screening tests must be confirmed by supplemental test (e.g. the Western blot [WB]�20 yet in England and Wales21 and in Africa,22, 23 HIV infection is diagnosed without a �confirmatory� Western blot test.
No proof for the existence of HIV proteins The belief that certain proteins are those of a unique retrovirus HIV originate from data published in Science in 1983 by Montagnier and his colleagues from the Pasteur Institute24 and in 1984 by Gallo and his colleagues at the National Institutes for Health in the United States.25 However, a thorough review of these papers26-31 fails to reveal any evidence that proteins attributed to a novel retrovirus HIV were obtained from purified retroviral-like particles proven infectious, that is, from a retrovirus. It is important to stress that proof of infectivity, that is, the ability to replicate, is a sine qua non in proving retroviral isolation. This is because retrovirologists have cautioned "Release of virus-like particles morphologically and biochemically resembling type-C virus [retrovirus] but apparently lacking the ability to replicate have been frequently observed from leukaemic tissue".32 That is, the demonstration of retroviral-like particles with reverse transcriptase activity is not proof for the existence of such viruses.33-35 The material which Montagnier and Gallo designated as �purified virus� was obtained from mitogenically stimulated cell cultures/co-cultures derived from AIDS patients. Aliquots of supernatants from these cultures were centrifuged through a sucrose density gradient and the material which banded at 1.16 gm/ml was claimed to be the �purified virus�. A decade earlier, a group of several leading retrovirologists stressed that proof of retroviral isolation requires examination of banded material with the electron microscope in order to establish �virus count, morphology and purity�. Proof of purity requires that the 1.16 gm/ml band contains nothing else but retroviral-like particles with "No apparent differences in physical appearances",33, 36 that is, all the particles must have the morphology of retroviral particles. However, neither Montagnier nor Gallo published electron micrographs (EM) of their �purified� material. Thus, neither Montagnier nor Gallo in 1983/84, nor at any time since, demonstrated that the material they claimed to be �purified HIV� consists of any particles, be they retroviral, viral or of any other morphologies, pure or impure. Both groups did publish EM of their �unpurified� cell cultures which demonstrated a small number of particles claimed to be the novel retrovirus HIV but these particles were not purified and proven infectious. Indeed these particles were claimed to belong to a retroviral genus known as C-type Oncoviruses, a genus taxonomically distinct from Lentiviruses in which HIV is classified.24, 25 Neither these EM nor any published since, confirm the existence of �HIV� particles bearing all the principal morphological characteristics of retroviruses.37 Subsequent studies reveal that T‑cell and monocyte "HIV infected cultures" contain, in addition to particles with the morphologies attributed to HIV, many other "viral particles" unlike any of the "HIV particles".37-40 Non‑HIV infected H9 cells, from which most of the published EM originate, as well as other cells used for "HIV isolation", CEM, C8166, EBV transformed B‑cells, and cord blood lymphocytes, express budding virus‑like particles albeit bearing somewhat different morphologies from particles deemed to be HIV.41 These data raise questions not only in regard to the origin and role of the "non‑HIV particles", but also the "HIV particles", and as to which, if any of these particles, band at 1.16 gm/ml. Most importantly, in the only EM study either in vivo or in vitro in which suitable controls were used and in which extensive blind examination of controls and test material was performed, particles indistinguishable from HIV were found in 18/20 (90%) of AIDS as well as in 13/15 (88%) of non-AIDS related lymph lymphadenopathies.42
The existence of HIV proteins is not predicated on an origin from retrovirus particles (RVP) or even material known to contain retrovirus-like particles. Instead it is predicated on reactions between a few of the many proteins present in the material banding at 1.16 gm/ml band, regarded as the �purified virus�, and antibodies present in the sera of AIDS patients. However, as argued above, from an antibody/antigen reaction it is impossible to define the origin of a protein or conclude that reacting antibodies arise as a result of specific exposure to that protein. Subsequently, the proteins attributed to �HIV� have been found in non-HIV-infected tissues and tissues of healthy individuals at no risk of AIDS. These include the p18,8, 10, 43, 44 p24,43-48 p32,49-51 p41,24, 49, 50, 52-56 and p120/160 proteins.43, 57, 58 From the beginning Montagnier has consistently stated that p41 is not an HIV protein but the ubiquitous cellular protein actin.24, 52 In 1989 Pinter et al57 demonstrated that the p120 and p160 proteins in the HIV Western blot strips are not distinct proteins but oligomers of p41 and that �confusion over the identification of these bands� has resulted in �incorrect conclusions�.58 Since the criteria for a positive Western blot in Africa59 requires reactivity with any two of p41, p120 or p160 regardless of any other bands, we may conclude that Africans are infected with HIV if they possess antibodies which react with one of their own proteins.
In 1997, fourteen years after its discovery, two events occurred which further compromised the notion of the existence of proteins specific to a novel, unique retrovirus. In March that year the first published EM of �purified virus� appeared in Virology. That is, EM of the sucrose density gradient banded material from which proteins and nucleic acids are obtained for use as diagnostic reagents, research and vaccine development.60, 61 These micrographs reveal �purified virus� as cellular, microvesicles interspersed with small numbers of particles possessing some but not all the morphological features of RVP. In the micrographs published by Bess and his colleagues �HIV� has a diameter of 234 nM, twice that of any other published �HIV� particle or known retrovirus. More importantly, according to Bess, comparison of the protein electrophoretic patterns of �infected� material banded through the sucrose density gradient with similarly banded �non-infected� material permits a �conclusion from gel electrophoresis patterns that there are only quantitative differences between HIV and microvesicles�. In other words, all the proteins present in the �purified virus� are also present in the cellular microvesicles. That is, the �HIV� particles and proteins are nothing but microvesicles and cellular proteins respectively. Other experiments conducted by the same group demonstrated that immunisation of monkeys with human cells cultured with and without �simian immunodeficiency virus�, produced an identical pattern of antibody response.50 This confirmed the 1991 experiments of Stott that 2 of 4 macaques immunised with non-infected human cells were protected from infection when challenged with �SIV� propagated in the same human cells62 and that protection �is correlated with the levels of antibody response to cellular antigens in the human cells from which the virus immunogen was grown�.63 For obvious reasons similar human, HIV data are not available.
In July 1997 Montagnier was interviewed en camera at the Pasteur Institute by the French investigative journalist Djamel Tahi and asked why he did not publish electron micrographs of �purified virus� in his seminal, 1983 Science paper. Montagnier�s response is astounding. He replied that, despite �a Roman effort� by his group, the particles observed in electron micrographs of the 1.16 gm/ml band, the �purified virus�, �did not have the morphology typical of retroviruses�. Pressed further he did not accept that Gallo�s laboratory had purified HIV. When asked �Gallo did it?� he replied, �Gallo?�I don�t know if he really purified. I don�t believe so�. Yet during the same interview Montagnier also asserted �analysis of the proteins of a virus demands mass production and purification� but �I repeat, we did not purify�.64
No proof of antibody specificity for HIV infection As the examples of infectious mononucleosis and syphilis amply illustrate, what is fundamental to an antibody test is not the origin and nature of the antigens employed but the specificity of the antibody reactivity induced by whatever agent the clinician is endeavouring to demonstrate. By definition, specificity is the proportion of HIV free individuals having a negative antibody test. In a hundred percent specific test every non-HIV-infected individual tests negative denoting a false-positive rate of zero (Figure 165). Thus, and in the words of the World Health Organization authorities,66 proving specificity rests exclusively upon a means of distinguishing between individuals �truly HIV-infected� and those �truly HIV-free�. Obviously the means to achieve this distinction cannot be the antigen/antibody reaction as this would amount to an antibody test acting as its own gold standard. Such a practice would, for example, be no different from endorsing pulmonary ventilation/perfusion scanning or exercise ECG stress testing by repetitive testing rather than by comparison with pulmonary or coronary angiography. A gold standard must be independent of whatever is being appraised and, in the case of HIV infection, can only be HIV itself, that is, HIV isolation/purification performed on the same individuals at the same time as the antibody tests. However, such a gold standard has never been applied. This is because to date there is no proof that any scientist, laboratory or institution has achieved HIV isolation/purification. Thus at present it remains impossible to define the specificity of the HIV antibody tests, a fact affirmed by the disclaimer repeatedly present in the Abbott Laboratories packet insert, �At present, there is no recognized standard for establishing the presence or absence of antibodies to HIV-1 and HIV-2 in human blood�.67, 68
Specificity = True negatives/(False positives+ True negatives) = (1-False positive rate)
Figure 1
Evidence presented as proof of specificity Despite lack of scientific evidence, the CDC and WHO simply assert that the HIV antibody tests are �extraordinarily accurate� in terms of sensitivity and specificity.69 Examination of the data published by HIV experts, institutions and biotechnology companies reveal methodologies which ignore validation against an independent gold standard. The examples below illustrate processes which use as a gold standard either the clinical AID syndrome, or �reference� sera, that is, sera defined to contain or not contain �HIV� antibodies on the basis of the presence or absence of AIDS. In the final analysis all methodologies amount to using the clinical syndrome or antibody reactivity acting as its own gold standard.
1. In 1985 Gallo and his colleagues defined individuals �truly HIV-infected� or �truly HIV-free� using the presence or absence of AIDS as their gold standard. They reported the sensitivity and specificity of the ELISA as 97.73% and 97.67% respectively.70 Clearly, it is scientifically impossible to draw conclusions regarding the existence of HIV on the basis of clinical data. One cannot use the AID clinical syndrome to define HIV status because this would require prior proof that all AIDS patients are �truly HIV-infected� and all non-AIDS individuals are �truly HIV-free�. Neither proposition is tenable because (a) patients with particular diseases are defined as AIDS on the basis of an antibody test; (b) since the vast majority of individuals with positive tests do not have the clinical syndrome, all positive tests in such individuals, be they sick or healthy, are false positives. Another significant flaw is restricting the choice of the non-AIDS �gold standard� to individuals who are healthy, such as blood donors or applicants for military service. Indeed, if generally applied this practice would prove almost any test highly specific. The net effect is to exhibit the test to a relatively narrow range of antibodies at the expense of ignoring the increased probability of cross reacting, �non-HIV� antibodies brought about by a large, heterogeneous population of antibodies typically present in sick individuals including those who are immune deficient and who may have diseases similar to but not AIDS. In fact, in a unique, largely ignored and never followed up study conducted at 26 US hospitals in 1990, amongst 89,547 anonymous blood specimens meticulously chosen to avoid patients at even meagre risk of AIDS, between 0.7% to 21.7% of men and 0-7.8% of women aged 25-44 years were found to be HIV infected on the basis of an ELISA and Western blot testing protocol.71
2. In the case of Abbott Laboratories �Sensitivity for HIV-1 antibodies was computed based on the clinical diagnosis of AIDS�Specificity is based on assay of blood donations from random donors�Specificity based on an assumed zero prevalence of antibody to HIV-1 and/or HIV-2 in random donors (17037 out of 17054) is estimated to be 99.90%� but �in these calculations one sample of the eighteen total repeatedly reactive specimens was confirmed by Western blot and has been excluded�.68 Significantly Abbott defines specificity on the basis of �antibody to HIV-1�, not HIV infection. This begs the question since one cannot presume �antibody to HIV-1� without prior proof of the specificity of the reactions. In addition, if �Specificity [is] based on an assumed zero prevalence of antibody to HIV-1�in random donors� then no seropositive blood donor can be infected with HIV. The practice of removing samples after Western blot testing negates the premises of the experiment as well as confirming use of one antibody test as the gold standard for another.
3. The Australian National Serology Laboratory (NRL) adopt a procedure exemplified by �Specificity monitoring of anti-HIV testing� at the Australian Red Cross Blood Service.72 Between 1985-2000 �An average of 855 329 specimens were tested for anti-HIV annually�. The NRL then calculates �The proportion of initially reactive [ELISA] results (after confirmed positives removed)��and repeatedly reactive [ELISA] (biologically falsely reactive) results�. The latter are those reactive on ELISA which are not �confirmed positives� using the Western blot. These data are reported as �Monitoring of false reactor rates (specificity)�. This can be criticised on several grounds: (a) using one antibody test (the Western blot) as the gold standard for another (ELISA); (b) failure to apply even this �gold standard� to all samples. That is, those non-reactive either initially or after a second ELISA, or those not Western blot positive after two reactive ELISAs. Even if this gold standard were de jure, without these data it is impossible to determine the number �truly HIV-free� required by definition to calculate the specificity of an antibody test.
4. Burke and his colleagues, when the former was Director, US Military HIV/AIDS Research Program, Division of Retrovirology, Walter Reed Army Institute of Research, determined specificity in a healthy population of 135,187 applicants for US military service. All applicants were screened with an initial ELISA and all reactive ELISAs were repeated in duplicate. Then an initial Western blot was performed and, if diagnostic or reactive, a second Western blot was performed on another fresh blood specimen. A positive Western blot was diagnosed if and only if the first and second serum samples were diagnostic. All the diagnostic Western blot samples were then assayed with four additional antibody tests. A Western blot was considered "true positive if all four assays on all available serum samples from an applicant were reactive and diagnostic", but was considered "false positive if all four assays on all available serum samples from an applicant were non‑reactive, non‑diagnostic or both". From the 135,187 applicants, there were 16 positive tests. In one of these, the serum was unavailable for further testing and one applicant declined to provide a second sample. Serum from 27 of the 29 samples from the 15 applicants found positive were tested by the four other antibody tests. Fourteen samples were found positive by all four assays and all four were negative for one applicant. From this Burke and his associates calculated his test procedure "has a specificity of roughly 99.9993%".73
Some of the many problems28 with this method are: (a) Burke�s and his colleagues define an arbitrary number (eight) of sequential antibody tests as the gold standard for being �truly HIV-infected�; (b) this de facto �gold standard� was applied to only 15 applicants again making it impossible to calculate specificity even by Burke�s method; (c) Burke�s premises are quite the opposite to those of Gallo and Abbott Laboratories where positive tests in healthy individuals are regarded as false positives.
5. For unknown reasons HIV experts also define HIV isolation on the basis of an antigen/antibody reaction. That is, a reaction between a laboratory produced antibody directed against a p24 protein and cell culture material. Even if one accepts this as proof of being �truly HIV-infected�, typical rates of �HIV isolation� are substantially less than seroprevalence rates.46 For example, the WHO analysed 224 specimens collected in Brazil, Rwanda, Thailand and Uganda from asymptomatic "HIV positive" individuals. Serostatus was first confirmed in the country of origin and then at the "centralized laboratories responsible for confirming serology, virus isolation, virus expression, and distribution of reagents (George-Speyer-Hans Chemotherapentisches Forschunginstitut (GSH) in Frankfurt, Germany; National Institute for Biological Standards and Control (NIBSC) in London, United Kingdom, and DAIDS/NIAID in Bethesda, Maryland, United States". Using the anti-p24-antibody method as �isolation�, "of a total of 224 virus cultures, 83 were positive (isolation rate=37%)"..74 This is virtually the same rate as Gallo�s original report a decade earlier where he and his colleagues claimed to have isolated HIV from 26/72 (36%) of AIDS patients.25 Accordingly, at least 60% of positive antibody tests occur in individuals from whom HIV cannot be isolated and thus are false positives.
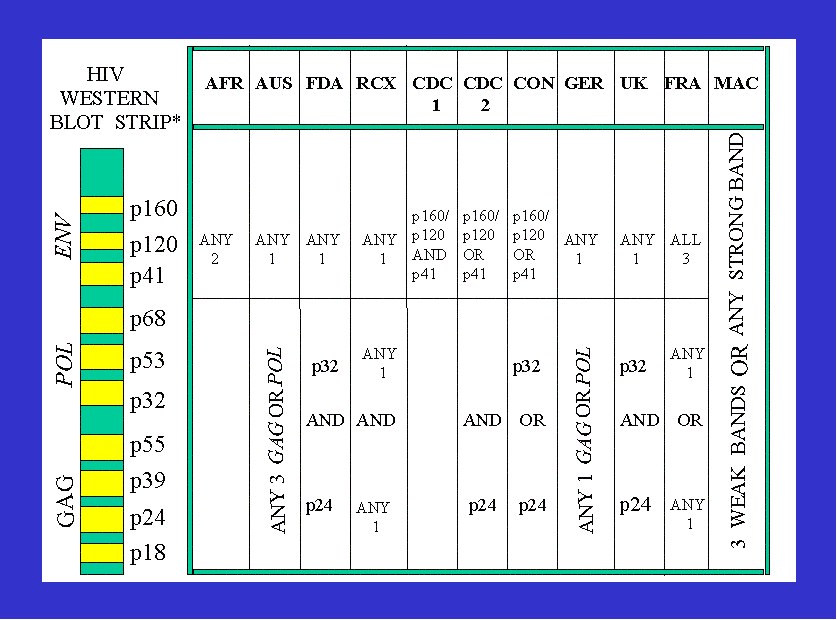
The HIV Western blot If there are such entities as HIV proteins capable of inducing specific antibody reactivity, one would expect reactivity with even one protein to constitute proof of infection. Indeed, prior to 1987, one "HIV specific" WB band was considered in this light. However, since 15%-25% of healthy, no risk individuals were found to have "HIV specific" WB bands,75, 76 it became necessary to redefine a positive WB by adding extra and selecting particular bands, otherwise at least one in every seven people would be diagnosed HIV infected. (Notwithstanding, in the Multicenter AIDS cohort study of 5000 gay men, one �strong� band remained proof of HIV infection up until 1990.77). Even if we ignore every problem detailed above it is still impossible to justify the claim that the HIV Western blot is �extraordinarily� specific (Table 1) because the criteria for a positive test vary between laboratories, institutions and countries. This gives rise to the anomaly where, for example, an individual positive in New York City on the CDC criteria would not be positive in Sydney, Australia. Or an Australian positive with p41, p32, p24 and p18 bands would not be positive in Africa. Or an African positive with a p41 and p120 band would not be positive in Australia, parts of the US or Europe. Confusion over antibody reactivity is confirmed in diagnostic laboratory manuals. The Genelabs Diagnostic HIV BLOT 2.2 Western blot Assay Instruction Manual advises, �Specific guidelines for interpretation may differ depending on the local policies, GENELABS recommends following the accepted policy to be in accordance with local regulations�. This is followed by seven different criteria for defining a positive Western blot issued by �different international regulatory bodies�. However, Genelabs also append, �We recommend the following guidelines for the interpretation of the Genelabs Diagnostic HIV BLOT 2.2� and list an eighth set of criteria for a positive Western blot.78 One wonders how �different international regulatory bodies� or �local policies�, and not the presumed pathogen, determine patterns of antibody reactivity that prove a viral infection. And why such patterns are geographically determined. Manufacturer Bio-Rad advises �Each laboratory performing Western blot testing should develop its own criteria for band interpretation. Alternatively, band interpretation may be left to the clinician" (Bio-Rad Laboratory Manual 1993).
In Australia the criteria for a positive Western blot consist of set combinations of four reactive bands.79 This means that one, two or three bands must be attributed to �non-HIV� antibodies47, 74, 80-88 (yet in others parts of the world some combinations of two or three bands would be caused by �HIV� antibodies). This raises the question, what is the scientific basis of the NRL criteria for an �Australian� positive Western blot and the claim it is virtually 100% specific for HIV infection? In other words, what scientific data prove that four or more, including all ten bands, cannot also be caused by �non-HIV� antibodies bearing no relationship to a retroviral infection? Especially given that AIDS patients are hypergammaglobulinaemic, all antibodies possess propensities for cross-reactivity and hypergammaglobulinaemia predicts seropositivity.89 In 199490 we sought an answer from the NRL but the response91 did not address the issue of proving specificity and subsequent attempts to obtain a definitive answer were unrewarding. However, this year the NRL informed the authors �It is most important that the conditions of the blot as recommended by manufacturers are followed. The National Serology Reference Laboratory, Australia advises this and the Therapeutic Goods Administration insists upon it�individual tests are interpreted according to manufacturer's directions�. The latter appears to contradict Genelabs which �recommends following the accepted policy to be in accordance with local regulations� (Elizabeth Dax, Director, NRL, personal communication).
Some of the best known experts on HIV antibody testing acknowledge that the Western blot has many shortcomings. According to Philip Mortimer, from the Central Public Health Laboratory, Virus Reference Laboratory, London, "The shortcomings of the western blot test would have been identified sooner if it had been evaluated as a condition of licensing and release on to national markets. Instead, this test has been used as the "gold standard" for other assays and the possibility that it might itself be inaccurate has largely been ignored�Western blot detection of HIV antibodies began as, and should have remained, a research tool".92 According to Dax, "Western blots present a series of disadvantages including their high cost, the lack of standardization in their production and their subjective reading".93 Yet the Western blot, whose inaccuracy has been �largely ignored�, is used as a gold standard for other antibody as well as PCR tests,94 not to mention itself,73 and is considered the �supplemental�, �confirmatory� proof of HIV infection.
If not a retrovirus what is the cause of antibody reactivity? The first possibility is that reactivity reflects specific or cross-reactions between cellular antigens and autoantibodies. This interpretation is supported by the finding of a plethora of antibodies in the sera of AIDS patients which react with �self� components. A large and growing number have been demonstrated including rheumatoid factor, anti‑cardiolipin, anti‑nuclear factor, anti‑cellular, anti‑platelet, anti‑red cell, anti‑actin, anti‑DNA, anti‑tubulin, anti‑thyroglobulin, anti‑albumin, anti‑myosin, anti‑thymosin, anti-lactoferrin, anti-TNF-α, anti-beta-2 glycoprotein I, anti-prothrombin, anti-neutrophil cytoplasmic, anti-ssDNA, anti-RNA, anti-histones, anti-nuclear antigen SS-A, anti-mitochondrial, anti-reticulin, anti-smooth muscle, anti-gut epithelial cell, anti-lymphocytic ganglioside, anti-Fab, anti-protein S, anti-brain proteins, anti-synthetic peptides of ubiquitinated histone H2A, anit-Sm-D antigen, anti-U1-A RNP antigen, anti-60 kD SSA/Ro antigen, anti-histone H1 and anti-histone H2B antibodies. Significantly, anti‑lymphocyte auto‑antibodies occur in 87% of seropositive individuals and their levels correlate with clinical status28, 29 (additional references available on request).
The other possibilities are that �HIV� antibodies are cross-reacting and non-specifically induced antibodies which arise because individuals belonging to the AIDS risk groups are exposed to a multitude of �non-HIV� antigens including those of many pathogenic agents as well as inanimate biological material. The former include viruses, bacteria, mycobacteria and fungi and the latter proteins and drugs which may induce the synthesis of antibodies. In Africa Kashala, Essex and their colleagues have shown that antibodies to carbohydrate-containing antigens such as lipoarabinomannan and phenolic glycolipid which constitute the cell wall of Mycobacterium leprae, a bacterium which �shares several antigenic determinants with other mycobacterial species� cause �significant cross-reactivities with HIV-1 pol and gag [p32, p55, p68, p24, p18] proteins�. This led the authors to warn that among leprosy patients and their contacts there is a �very high rate of HIV-1 false-positive ELISA and WB results�, that �ELISA and WB results should be interpreted with caution when screening individuals infected with M. tuberculosis or other mycobacterial species�, and furthermore that �ELISA and WB may not be sufficient for HIV diagnosis in AIDS-endemic areas of central Africa where the prevalence of mycobacterial diseases is quite high�.95 In fact "Mycobacterium tuberculosis (MTB) disease is an extremely important life-threatening bacterial disease with 8 million new cases and 3 million deaths reported world-wide each year to the World Health Organization; the vast majority of these cases are in developing countries�The World Health Organization has estimated that 5.6 million worldwide and 80,000 people in the United States are co-infected with HIV and MTB".96 Furthermore, the developing world "bears more than 90% of the global burden of HIV infection" and "Tuberculosis (TB) is the leading cause of death worldwide among people with HIV�.97 No less an authority on AIDS in Africa than De Cock acknowledges that TB has been present in an epidemic proportion in developing countries long before the AIDS era.98
Not only mycobacteria (M. leprae, M. tuberculosis, M. avium-intracellulare) but also the walls of fungi (Candida albicans, Cryptococcus neoformans, Coccidioides immitis, Histoplasma capsulatum, including Pneumocystis carinii), contain carbohydrate mannans. "The immunochemical determinants of the antigenic factors of Candida albicans display a high identity with the glycoprotein (gp) 120 of HIV-1: they contain α(1→2) and α(1→3) linked mannose terminal residues".99 "One half of the molecular weight of gp120 is represented by oligomannosidic oligosaccharides...Polyclonal antibodies to mannan from yeast also recognize the carbohydrate structure of gp120 of the AIDS virus".100 Antibodies to the mannans of Candida albicans "block infection of H9 cells by HIV-1" as well as the binding of lectins to gp120.99 Recognition of gp120 by antibodies to a synthetic peptide of the same antigen was "partially abolished if it was absorbed with the total polysaccharide fraction of C. albicans" while the antigen recognition by antibodies to "gp120 from human T cell lymphotropic virus type IIIB [HIV]", "was totally blocked". From these data the authors concluded: "These results indicate that mannan residues of C. albicans can serve as antigens to raise neutralizing antibodies against HIV infection�.99 One hundred per cent of AIDS patients (even those with �no Candida clinically�) have C. albicans antibodies.101 Significantly, PCP, candidiasis, cryptococcosis, coccidioidomycosis, histoplamosis, tuberculosis or Mycobacterium avium-intracellular constitute the opportunistic infections present in 88% of AIDS cases diagnosed between 1988 and 1992.102
How may a physician judge the significance of a positive �HIV� antibody test?Although AIDS began to decline in 1987,103, 104 this trend was countered by the addition of more and more diseases including, in 1993, laboratory abnormalities in the absence of diseases,105 to each revision (1985, 1987, 1993 and 2000) of the original, 1982 CDC definition. At the same time antibody testing �evolved� such that the earliest assays which used �purified HIV lysates (1st generation), and often lacked sensitivity and specificity� were replaced by �Improved assays based on recombinant proteins and/or synthetic peptides�(2nd generation)� followed by �The so-called 3rd generation or sandwich ELISAs�.23 In other words, antibody tests based on �purified virus� antigens resulted in an inferior correlation with the clinical syndrome than the engineered �Improved assays� with their �recombinant proteins and/or synthetic peptides� which replaced them. Thus, by �refining� the test antigens, by altering the criteria for a positive test as well as the definition of AIDS, and by restricting testing largely to the risk groups, HIV experts have sought and obtained a high degree of correlation between the presence or absence of �HIV� antibodies and the presence or absence of the clinical syndrome.
Notwithstanding such partiality, (and also ignoring confounders such as psychological, behavioural and treatment factors which may prove injurious to health), there is no doubt, at least in the risk groups, that a positive antibody test does predict an increased likelihood of developing diseases including both AIDS indictor and non-AIDS diseases,106 and dying prematurely. Since there is no proof that a positive �HIV� antibody test is caused by a novel retroviral infection, the underlying reason for the correlation must be sought elsewhere. The explanation which accords with both non-specificity and clinical relevance is that the antibody tests are similar to measurements of the erythrocyte sedimentation rate (ESR). An elevated ESR "is a measure of the presence and intensity of morbid processes within the body" and, like a positive "HIV" antibody test, also has the capacity to predict "a likelihood of death within the next several years far above" a normal ESR.107 A common cause of elevated ESR is infection and other causes include malignancy, collagen vascular diseases, rheumatic heart disease, and other chronic disease states, including humans with a positive HIV antibody test. Even asymptomatic, non-anaemic HIV positive individuals may have an increased ESR108 and the test may be predictive for disease progression.109 In HIV positive children a correlation exists between seropositivity, hypergammaglobulinaemia and elevated ESR.110 As far back as 1988 researchers from the Institut National de Transfusion Sanguine, Paris, France, found that "An increased ESR in HIV-seropositive subjects seems to constitute a predictive marker of progression towards AIDS before the decrease of the CD4 count".111 In other words the ESR is a better predictive marker for AIDS than decrease in the CD4 cell count although the latter is said to be the cause of the syndrome. One important factor which affects the ESR is the size of the red cells, especially Rouleaux formation, where the red blood cells clump together. Rouleaux formation may result from changes in the negative change of red cells, caused by "the dielectric effect of proteins in the surrounding plasma", especially by "fibrinogen, immunoglobulins, and other acute-phase reaction proteins" and their increased levels in some disease states.107 Diseases such as tuberculosis and AIDS are not caused by red blood cell clumping induced by "the dielectric effect of proteins" but the fact this can be demonstrated and measured in vitro is of great utility in clinical practice. This applies both to diagnosis and monitoring the course of the disease including response to treatment. Thus it appears that in the mid 1980s laboratory scientists serendipitously discovered an ESR-like test but, under great pressure and in great haste to find the cause of a new and alarming syndrome, reported it as HIV infection112 and recommended its general use as a diagnostic tool for this purpose. Unfortunately, the historical caveat provided by HL23V, the world�s �first� human retrovirus, enthusiastically proposed in the mid 1970s as a �real possibility� for an infectious cause of human leukaemia,113 was either unheeded or forgotten. This �retrovirus�, discovered in 1975114 and whose short existence was also predicated on antibody reactivity, suffered a precipitous demise in 1980 when such reactivity was proven non-specific.115, 116
Given that the "HIV" proteins are normal cellular proteins, or proteins with new antigenic epitopes, or newly induced cellular proteins, and that individuals who test positive have high levels of autoantibodies and/or antibodies to many non-HIV antigens, all or some of which may cross-react with cellular proteins, "HIV" seropositivity, like the ESR, may represent nothing more than a non-specific indicator of altered homeostasis connoting a propensity to or the presence of particular diseases. Whether or not a positive antibody test is predictive outside the risk groups remains to be proven. As long as the present interpretation of a positive test is accepted, this may never be ascertained since knowledge of seropositivity by both patient and physician attracts multiple confounding factors virtually impossible to eliminate in a definitive analysis.
Conclusion The parameters of an antibody test for HIV infection can be defined only by reference to HIV isolation/purification. To date this has not been reported and use of the clinical syndrome and �reference� sera as de facto gold standards is scientifically inadmissible as they amount to using an antibody test as its own gold standard. Thus there is no scientific basis to the claim that the HIV antibody tests are �extraordinarily accurate� for diagnosing HIV infection. In fact the weight of evidence favours the opposite conclusion because (a) there is no proof that a retrovirus HIV has been isolated/purified; (b) test antigens are obtained from cellular material lacking particles taxonomically consistent with RPV; (c) there is proof that the �HIV� antigens are cellular proteins and AIDS patients generate numerous autoantibodies; (d) numerous data confirm reactivity caused by �non-HIV� antibodies in both animals and humans and this is particularly likely given the hypergammaglobulinaemia typical of the clinical AID syndrome; (e) depending on where an individual is tested, one and the same serum reaction is interpreted as being caused by �non-HIV� antibodies (�indeterminate�) or by �HIV� antibodies (�positive�).
The data presented in this paper call into question the �overwhelming evidence� said to substantiate a positive antibody test as proof of HIV infection. Public, scientific debate on the role of �non-HIV� and non-infectious factors in the genesis of a positive antibody test is long overdue. 117-122
VARIATION IN GLOBAL CRITERIA FOR DEFINING A POSITIVE WESTERN BLOT
AFR=AFRICA;1 AUS=AUSTRALIA;2 FDA=US FOOD AND DRUG ADMINISTRATION;3 RCX=US RED CROSS;3 CDC=US CENTER FOR DISEASE CONTROL;3 CON=US CONSORTIUM FOR RETROVIRUS SEROLOGY STANDARDIZATION;3 GER=GERMANY; UK=UNITED KINGDOM; FRA=FRANCE; MACS= US MULTICENTER AIDS COHORT STUDY 1983-1992. * Bands not in electrophoretic order
NOTES: I. �The Association of Public Health Laboratories now recommends that patients who have minimal positive results on the WB, eg p24 and gp160 only, or gp41 and gp160 only, be told that these patterns have been seen in persons who are not infected with HIV and that follow-up testing is required to determine actual infective status�.4 II. In February 1993 the US Food and Drug Administration relaxed their criteria in order to �reduce the number of HIV-1 seroindeterminate Western blot interpretations�, that is, to increase the number of HIV positive individuals.5
1. WHO. (1990). Acquired Immunodeficiency Syndrome (AIDS). Proposed criteria for interpreting results from Western blot assays for HIV-1, HIV-2 and HTLV-I/HTLV-II. Weekly Epidemiological Record 65:281-298. 2. Healy DS, Maskill WJ, Howard TS, et al. (1992). HIV-1 Western blot: development and assessment of testing to resolve indeterminate reactivity. AIDS 6:629-633. 3. Lundberg GD. (1988). Serological Diagnosis of Human Immunodeficiency Virus Infection by Western blot Testing. Journal of the American Medical Association 260:674-679. (Data presented in this paper reveal that when the FDA criteria are used to interpret the HIV Western blot less than 50% of US AIDS patients are HIV positive whereas 10% of persons not at risk of AIDS are also positive). 4. Mylonakis E, Paliou M, Greenbough TC, Flaningan TP, Letvin NL, Rich JD. Report of a false-positive HIV test result and the potential use of additional tests in establishing HIV serostatus. Archives of Internal Medicine 2000;160:2386-8. 5. Keinman S, Busch MP, Hall L, et al. (1998). False-positive HIV-1 test results in a low -risk screening setting of voluntary blood donation. Journal of the American Medical Association 280:1080-1083.
NOTE: Each horizontal band on the left represents a protein with which an antibody can react. Serum from a patient is added to a strip and the strip developed. Where there have been antibodies reacting a dark band occurs. The number and location determining a positive test, for the same virus, varies all over the world. Even today there are still no internationally agreed criteria as to what constitutes a positive WB. This gives rise to the bizarre situation where, for example, an individual positive in New York City on the CDC criteria would not be positive in Sydney, Australia. Or an Australian positive with p41, p32, p24 and p18 bands would not be positive in Africa. Or an African positive with a p41 and p120 band would not be positive in Australia, parts of the US or Europe. Confusion over antibody reactivity is confirmed in diagnostic laboratory manuals. The Genelabs Diagnostic HIV BLOT 2.2 Western blot Assay Instruction Manual advises, �Specific guidelines for interpretation may differ depending on the local policies, GENELABS recommends following the accepted policy to be in accordance with local regulations�. This is followed by seven different criteria for defining a positive Western blot issued by �different international regulatory bodies�. Genelabs also append, �We recommend the following guidelines for the interpretation of the Genelabs Diagnostic HIV BLOT 2.2� and list an eighth set of criteria for a positive Western blot. This means that �different international regulatory bodies� or �local policies�, and not the presumed pathogen determine patterns of antibody reactivity said to prove a retroviral infection. Manufacturer Bio-Rad advises �Each laboratory performing Western blot testing should develop its own criteria for band interpretation. Alternatively, band interpretation may be left to the clinician" (Bio-Rad Laboratory Manual 1993). REFERENCES
1. von Behring E, Kitazato S. Ueber das Zustandekommen der Diphtherie-Immunit�t und der Tetanus-Immunit�t bei Thieren. Deutsche Medizinische Wochenschrift 1890;16:1113-1114. 2. M�rner PtCKAH. 1901. Emil Adolf von Behring, Nobel Presentation Address. www.nobel.se/medicine/laureates/1901/press.html 3. Nossal GJV. Antibodies and Immunity. Harmondsworth, UK: Penguin Books Ltd; 1971. 4. Guilbert B, Fellous M, Avrameas S. HLA-DR-specific monoclonal antibodies cross-react with several self and nonself non-MHC molecules. Immunogenetics 1986;24:118-121. 5. Pontes de Carvalho LC. The faithfullness of the immunoglobulin molecule: can monoclonal antibodies ever be monospecific? Immunol. Today 1986;7:33. 6. Ternynck T, Avrameas S. Murine natural monoclonal antibodies: a study of their polyspecificities and their affinities. Immunol. Rev. 1986;94:99-112. 7. Owen M, Steward M. Antigen recognition. In: Roitt I, Brostoff J, Male D, editors. Immunology. 4th ed. London: Mosby; 1996. p. 7.1-7.12. 8. Chassagne J, Verelle P, Fonck Y, Legros M, Dionet C, Plagne R, et al. Detection of the lymphadenopathy-associated virus p18 in cells of patients with lymphoid diseases using a monoclonal antibody. Annales de l Institut Pasteur - Immunology 1986;137D:403-8. 9. Gonzalez-Quintial R, Baccala R, Alzari PM, Nahmias C, Mazza G, Fougereau M, et al. Poly(Glu60Ala30Tyr10) (GAT)-induced IgG monoclonal antibodies cross- react with various self and non-self antigens through the complementarity determining regions. Comparison with IgM monoclonal polyreactive natural antibodies. Europ. J. Immunol. 1990;20:2383-7. 10. Parravicini CL, Klatzmann D, Jaffray P, Costanzi G, Gluckman JC. Monoclonal antibodies to the human immunodeficiency virus p18 protein cross-react with normal human tissues. AIDS 1988;2:171-177. 11. Fauci AS, Lane HC. Human Immunodeficiency Virus (HIV) Disease: AIDS and Related Disorders. In: Isselbacher KJ, Braunwald E, Wilson JD, Martin JB, Fauci AS, Kasper DL, editors. Harrisons Principles of Internal Medicine. 13th ed. New York: McGraw-Hill Inc.; 1994. p. 1566-1618. 12. Berzofsky JA, Berkower IJ, Epstein SL. Antigen-Antibody Interactions and Monoclonal Antibodies. In: Paul WE, editor. Fundamental Immunology. 3rd ed. New York: Raven; 1993. p. 421-465. 13. Laal S, Samanich KM, Sonnenberg MG, Zolla-Pazner S, Phadtare JM, Belisle JT. Human humoral responses to antigens of Mycobacterium tuberculosis: immunodominance of high-molecular-mass antigens. Clin. Diagn. Lab. Immunol. 1997;4:49-56. 14. Baskar PV, Collins GD, Dorsey-Cooper BA, Pyle RS, Nagel JE, Dwyer D, et al. Serum antibodies to HIV-1 are produced post-measles virus infection: evidence for cross-reactivity with HLA. Clin. Exp. Immunol. 1998;111:251-6. 15. Marseille E, Kahn JG, Mmiro F, Guay L, Musoke P, Fowler MG, et al. Cost effectiveness of single-dose nevirapine regimen for mothers and babies to decrease vertical HIV-1 transmission in sub-Saharan Africa. Lancet 1999;354:803-9. 16. Colebunders RI, Greenberg A, Nguyen-Dinh P, Francis H, Kabote N, Izaley L, et al. Evaluation of a clinical case definition of AIDS in African children. AIDS 1987;1:151-3. 17. Dansette PM, Bonierbale E, Minoletti C, Beaune PH, Pessayre D, Mansuy D. Drug-induced immunocytotoxicity. Eur. J. Drug Metab. Pharmacokinet. 1998;23:443-451. 18. Roitt IM. Roitt's Essential Immunology. Ninth ed. London: Blackwell Science; 1997. 19. Crawford DH, Azim T. The use of the Epstein-Barr virus for the production of human monoclonal antibody secreting cell lines. In: Brown J, editor. Human monoclonal antibodies: Current techniques and future perspectives. Oxford: IRL Press Ltd; 1986. p. 1-6. 20. CDC. Sexually transmitted diseases treatment guidelines. Morb. Mortal. Wkly. Rep. 2002;51 (No. RR-6):7-10. www.cdc.gov/std/treatment/rr5106.pdf 21. Mortimer PP. Ten years of laboratory diagnosis of HIV: how accurate is it now? J. Antimicrob. Chemother. 1996;37:B. 27-32. 22. Katzenstein D. Vertical transmission of HIV in Africa: diagnostic testing and new interventions. HIV Clinical Trials 2000;1:51-7. 23. WHO. Operational characteristics of commercially available assays to detect antibodies to HIV-1 and/or HIV-2 in human sera. 1999; Report 11. www.who.int/mipfiles/2333/OperationalCharacteristics_HIVReport11_MIP2001.pdf 24. Barr�-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983;220:868-71. 25. Gallo RC, Salahuddin SZ, Popovic M, Shearer GM, Kaplan M, Haynes BF, et al. Frequent Detection and Isolation of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and at Risk for AIDS. Science 1984;224:500-503. 26. Papadopulos E, Turner V. Presentation to the Presidential Panel AIDS Advisory Meeting 3 & 4 July Johannesberg, South Africa. 2000. www.theperthgroup.com/aids/pretoria2.doc 27. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Has Gallo proven the role of HIV in AIDS? Emerg. Med. [Australia] 1993;5:113-123. 28. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Is a positive Western blot proof of HIV infection? Bio/Technology 1993;11:696-707. 29. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Causer D. The Isolation of HIV: Has it really been achieved? Continuum 1996;4:1s-24s. www.virusmyth.com/aids/data/epreplypd.htm 30. Papadopulos-Eleopulos E. A critical analysis of the evidence for the existence of HIV and the HIV antibody tests: Lecture the XIIth International AIDS Conference, Geneva. 1998. www.virusmyth.com.aids/perthgroup/geneva 31. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM, Alfonso H, Page BAP, Causer D, et al. Mother to Child Transmission of HIV and its Prevention with ATZ and Nevirapine. Perth: The Perth Group; 2001. http://www.aidspanelreport.com/newperthpaper.htm 32. Gallo RC, Wong-Staal F, Reitz M, Gallagher RE, Miller N, Gillespie DH. Some evidence for infectious type-C virus in humans. In: Balimore D, Huang AS, Fox CF, editors. Animal Virology. New York: Academic Press Inc.; 1976. p. 385-405. 33. Toplin I. Tumor Virus Purification using Zonal Rotors. Spectra 1973:225-235. 34. Temin HM, Baltimore D. RNA-Directed DNA Synthesis and RNA Tumor Viruses. Adv. Virol. Res. 1972;17:129-186. 35. Bader JP. Reproduction of RNA Tumor Viruses. In: Fraenkel-Conrat H, Wagne RR, editors. Comprehensive Virology. New York: Plenum Press; 1975. p. 253-331. 36. Sinoussi F, Mendiola L, Chermann JC. Purification and partial differentiation of the particles of murine sarcoma virus (M. MSV) according to their sedimentation rates in sucrose density gradients. Spectra 1973;4:237-243. 37. Gelderblom HR, �zel M, Hausmann EHS, Winkel T, Pauli G, Koch MA. Fine Structure of Human Immunodeficiency Virus (HIV), Immunolocalization of Structural Proteins and Virus-Cell Relation. Micron Microscopica 1988;19:41-60. 38. Gendelman HE, Orenstein JM, Martin MA. Efficient Isolation and Propagation of Human Immunodeficiency Virus on Recombinant Colony-Stimulating Factor 1-Treated Monocytes. J. Exp. Med. 1988;167:1428-1441. 39. Lecatsas G, Taylor MB. Pleomorphism in HTLV-III, the AIDS virus. S. Afr. Med. J. 1986;69:793-794. 40. Hockley DJ, Wood RD, Jacobs JP. Electron Microscopy of Human Immunodeficiency Virus. J. Gen. Virol. 1988;69:2455-2469. 41. Dourmashkin RR, O'Toole CM, Bucher D, Oxford JS. The presence of budding virus-like particles in human lymphoid cells used for HIV cultivation. In: VIIth International Conference on AIDS; 1991; Florence; 1991. p. 122. 42. O'Hara CJ, Groopmen JE, Federman M. The Ultrastructural and Immunohistochemical Demonstration of Viral Particles in Lymph Nodes from Human Immunodeficiency Virus-Related Lymphadenopathy Syndromes. Hum. Pathol. 1988;19:545-549. 43. Faulk WP, Labarrere CA. HIV proteins in normal human placentae. Am. J. Reprod. Immunol. 1991;25:99-104. 44. Stricker RB, Abrams D, I,, Corash L. Target platelet antigen in homosexual men with immune thrombocytopenia. N. Engl. J. Med. 1985;313:1375-1380. 45. Agbalika F, Ferchal F, Garnier JP, Eugene M, Bedrossian J, Lagrange PH. False-positive HIV antigens related to emergence of a 25-30kD proteins detected in organ recipients. AIDS 1992;6:959-962. 46. Jackson JB, Kwok SY, Sninsky JJ, Hopsicker JS, Sannerud KJ, Rhame FS, et al. Human immunodeficiency virus type 1 detected in all seropositive symptomatic and asymptomatic individuals. J. Clin. Microbiol. 1990;28:16-9. 47. Mortimer P, Codd A, Connolly J, Craske J, Desselberger U, Eglin R, et al. Towards error free HIV diagnosis: notes on laboratory practice. Pub. Health Lab. Service Micrbiol. Digest 1992;9:61-64. 48. Vincent F, Belec L, Glotz D, Menoyo-Calonge V, Dubost A, Bariety J. False-positive neutralizable HIV antigens detected in organ transplant recipients. AIDS 1993;7:741-742. 49. Arthur LO, Bess JW, Sowder II RC, Beneviste RE, Mann DL, Chermann JC, et al. Cellular proteins bound to immunodeficiency viruses:implications for pathogenesis and vaccines. Science 1992;258:1935-1938. 50. Arthur LO, Bess JW, Jr., Urban RG, Strominger JL, Morton WR, Mann DL, et al. Macaques immunized with HLA-DR are protected from challenge with simian immunodeficiency virus. J. Virol. 1995;69:3117-24. 51. Henderson LE, Sowder R, Copeland TD. Direct Identification of Class II Histocompatibility DR Proteins in Preparations of Human T-Cell Lymphotropic Virus Type III. J. Virol. 1987;61:629-632. 52. Chamaret S, Squinazi F, Courtois Y, Montagnier L. Presence of anti-HIV antibodies in used syringes left out in public places, beaches or collected through exchange programs. In: XIth International Conference on AIDS; 1996; Vancouver; 1996. 53. Choudhury S, El-Farrash MA, Kuroda MJ, Harada S. Retention of HIV-1 inside infected MOLT-4 cells in association with adhesion-induced cytoskeleton reorganisation. AIDS 1996;10:363-368. 54. Pearce-Pratt R, Malamud D, Phillips DM. Role of cytoskeleton in cell-to-cell transmission of human immunodeficiency virus. J. Virol. 1994;68:2898-2905. 55. Orentas RJ, Hildreth JEK. Association of host cell surface adhesion receptors and other membrane proteins with HIV and SIV. AIDS Res. Hum. Retroviruses 1993;9:1157-1165. 56. Sasaki H, Nakamura M, Ohno T, Matsuda Y, Yuda Y, Nonomura Y. Myosin-actin interaction plays an important role in human immunodeficiency virus type 1 release from target cells. Proc. Natl. Acad. Sci. U S A 1995;92:2026-2030. 57. Pinter A, Honnen WJ, Tilley SA, Bona C, Zaghouani H, Gorny MK, et al. Oligomeric structure of gp41, the transmembrane protein of human immunodeficiency virus type 1. J. Virol. 1989;63:2674-9. 58. Zolla-Pazner S, Gorny MK, Honnen WJ. Reinterpretation of human immunodeficiency virus Western blot patterns. N. Engl. J. Med. 1989;320:1280-1281. 59. WHO. Acquired Immunodeficiency Syndrome (AIDS). Proposed criteria for interpreting results from Western blot assays for HIV-1, HIV-2 and HTLV-I/HTLV-II. Wkly. Epidem. Rec. 1990;65:281-298. 60. Bess JW, Gorelick RJ, Bosche WJ, Henderson LE, Arthur LO. Microvesicles are a source of contaminating cellular proteins found in purified HIV-1 preparations. Virol. 1997;230:134-144. 61. Gluschankof P, Mondor I, Gelderblom HR, Sattentau QJ. Cell membrane vesicles are a major contaminant of gradient-enriched human immunodeficiency virus type-1 preparations. Virol. 1997;230:125-133. 62. Stott EM. Anti-cell antibody in macaques. Nature 1991;353:393. 63. Chan WL, Rodgers A, Hancock RD, Taffs F, Kitchin P, Farrar G, et al. Protection in simian immunodeficiency virus-vaccinated monkeys correlates with anti-HLA class I antibody response. J. Exp. Med. 1992;176:1203-7. 64. Tahi D. Did Luc Montagnier discover HIV? Text of video interview with Professor Luc Montagnier at the Pasteur Institute July 18th 1997. Continuum 1998;5:30-34. www.virusmyth.com/aids/data/dtinterviewlm.htm 65. Griner PF, Mayewski RJ, Mushlin AI. Selection and interpretation of diagnostic tests and procedures. Ann. Int. Med. 1981;94:559-563. 66. National Institute of Allergy and Infectious Diseases. 2001. Focus on the HIV-AIDS Connection. www.niaid.nih.gov/newsroom/focuson/hiv00/default.htm 67. Laboratories A. Human Immunodeficiency Virus Type-1. Qualitative Enzyme Immunoassay for the Detection of Antibody to Human Immunodeficiency Virus Type-1 (HIV-1) in Human Serum or Plasma. Abbott Park. Illinois: United States of America: Abbott Laboratories, Diagnostics Division; 1988 1988. 68. Abbot Laboratories. Abbott Axsym system (HIV-1/HIV-2). 100 Abbott Park Rd. Abbott Park. Illinois: United States of America. 1998. 69. WHO. 1998. HIV Antibody ELISA Tests. www.who.int/pht/blood_safety/hivtable1.html 70. Weiss SH, Goedert JJ, Sarngadharan MG, Bodner AJ, Gallo RC, Blattner WA. Screening test for HTLV-III (AIDS agent) antibodies. Specificity, sensitivity, and applications. J. Am. Med. Assoc. 1985;253:221-5. 71. St. Louis ME, Rauch KJ, Peterson LR, Anderson JE, Schable CA, Dondero TJ. Seroprevalence rates of human immunodeficiency virus infection at sentinel hospitals in the United States. N. Engl. J. Med. 1990;323:213-218. 72. Hamilton CW, Dax EM. Quality and Laboratory Diagnosis of HIV Infection. Microbiol. Aust. 2001;22:23-25. 73. Burke DS, Brundage JF, Redfield RR, Damato JJ, Schable CA, Putman P, et al. Measurement of the false positive rate in a screening program for human immunodeficiency virus infections. N. Engl. J. Med. 1988;319:961-964. 74. WHO. HIV type 1 variation in World Health Organization-sponsored vaccine evaluation sites: genetic screening, sequence analysis, and preliminary biological characterization of selected viral strains. AIDS Res. Hum. Retroviruses 1994;10:1327-1343. 75. Lundberg GD. Serological diagnosis of human immunodeficiency virus infection by Western Blot testing. J. Am. Med. Assoc. 1988;260:674-679. 76. Wooldridge M. Commonwealth of Australia Minister for Health and Human Servies. Letter to Senator Christopher Ellison. 1997. 77. Phair J, Jacobson L, Detals R, Rinaldo C, Saah A, Schrager L, et al. Acquired Immune Deficiency Syndrome Occuring Within 5 Years of Infection with Human Immunodeficiency Virus Type-1: The Multicenter AIDS Cohort Study. J. Acquir. Immun. Defic. Syndr. 1992;5:490-496. 78. Genelabs Diagnostics Pty Ltd HIV BLOT 2.2 Instruction Manual. Singapore; 1999. 79. Healy DS, Maskill WJ, Howard TS, Armstrong VA, Bolton WV, Cooper GJ, et al. HIV-1 Western blot: development and assessment of testing to resolve indeterminate reactivity. AIDS 1992;6:629-633. 80. Genesca J, Jett BW, Epstein JS, Shih JWK, Hewlett IK, Alter HJ. What do Western Blot indeterminate patterns for Human Immunodeficiency Virus mean in EIA-negative blood donors? Lancet 1989;ii:1023-1025. 81. Ranki A, Johansson E, Krohn K. Interpretation of antibodies reacting solely with human retroviral core proteins. N. Engl. J. Med. 1988;318:448-9. 82. Belshe RB, Clements ML, Keefer MC, Graham BS, Corey L, Sposto R, et al. Interpreting HIV serodiagnostic test results in the 1990s: social risks of HIV vaccine studies in uninfected volunteers. Ann. Int. Med. 1994;121:584-589. 83. Bukrinsky MI, Chaplinskas SA, Syrtsev VA, Bravkilene LA, Philippov YV. Reactivity to gag- and env-related proteins in immunoblot assay is not necessarily indicative of HIV infection. AIDS 1988;2:405-6. 84. Kozhemiakin LA, Bondarenko IG. Genomic instability and AIDS. Biokhimiia 1992;57:1417-26. 85. Kion TA, Hoffmann GW. Anti-HIV and anti-anti-MHC antibodies in alloimmune and autoimmune mice. Science 1991;253:1138-1140. 86. Lang JM, Touraine JL, Trepo C, Choutet P, Kirstetter M, Falkenrodt A, et al. Randomised, double-blind, placebo-controlled trial of ditiocarb sodium ('Imuthiol') in human immunodeficiency virus infection. Lancet 1988;2:702-6. 87. Strandstrom HV, Higgins JR, Mossie K, Theilen GH. Studies with canine sera that contain antibodies which recognize human immunodeficiency virus structural proteins. Cancer Res. 1990;50:5628s-5630s. 88. Western Blot Assay HIV Blot 2.2. Genelabs Diagnostics Pty. Ltd. Singapore; 1996. 89. Brenner B, Schwartz S, Ben-Porath E, Tatarsky I, Varon D, Martinowitz U. The prevalence and interaction of human immunodeficiency virus and hepatitis B infections in Israeli hemophiliacs. Isr. J. Med. Sci. 1991;27:557-561. 90. Turner VF. The HIV Western blot. Med. J. Aust. 1994;160:807-808. 91. Dax E. The HIV Western blot: Reply to letter. Med. J. Aust. 1994;160:808. 92. Mortimer P. The fallability of HIV western blot. Lancet 1991;337:286-287. 93. Dax EM. Serological testing algorithms for the diagnosis of HIV infection. AIDS 1999;13:133-4. 94. Owens DK, Holodniy M, Garber AM, Scott J, Sonnad S, Moses L, et al. Polymerase chain reaction for the diagnosis of HIV infection in adults. A meta-analysis with recommendations for clinical practice and study design. Ann. Int. Med. 1996;124:803-15. 95. Kashala O, Marlink R, Ilunga M, Diese M, Gormus B, Xu K, et al. Infection with human immunodeficiency virus type 1 (HIV-1) and human T cell lymphotropic viruses among leprosy patients and contacts: correlation between HIV-1 cross-reactivity and antibodies to lipoarabinomannan. J. Infect. Dis. 1994;169:296-304. 96. Goletti D, Weissman D, Jackson RW, Graham NM, Vlahov D, Klein RS, et al. Effect of Mycobacterium tuberculosis on HIV replication. Role of immune activation. J. Immunol. 1996;157:1271-8. 97. Horton R. The 12th World AIDS Conference: a cautionary tale. Lancet 1998;352:122. 98. De Cock KM, Soro B, Coulibaly IM, Lucas SB. Tuberculosis and HIV infection in sub-Saharan Africa. J. Am. Med. Assoc. 1992;268:1581-7. 99. Muller WEG, Bachmann M, Weiler BE, Schroder HC, Uhlenbruck GU, Shinoda T, et al. Antibodies against defined carbohydrate structures of Candida albicans protect H9 cells against infection with human immunodeficiency virus-1 in vitro. J. Acquir. Immun. Defic. Syndr. 1991;4:694-703. 100. Muller WEG, Schroder HC, Reuter P, Maidhof A, Uhlenbruck G, Winkler I. Polyclonal antibodies to mannan from yeast also recognize the carbohydrate structure of gp120 of the AIDS virus: an approach to raise neutralizing antibodies to HIV-1 infection in vitro. AIDS 1990;4:159-162. 101. Matthews R, Smith D, Midgley J, Burnie J, Clark I, Connolly M, et al. Candida and AIDS: Evidence for protective antibody. Lancet 1988;ii:263-266. 102. Hu DJ, Fleming PL, Castro KG, Jones JL, Bush TJ, Hanson D, et al. How important is race/ethnicity as an indicator of risk for specific AIDS-defining conditions? J. acquir. immune def. syndr. hum. retrovirol. 1995;10:374-380. 103. Melbye M, Begtrup K, Rosenberg PS, Biggar RJ, Goedert J, Knudsen K, et al. Differences in susceptibility to AIDS development: A cohort study of Danish and American homosexual-bisexual men, 1981-1995. J. acquir. immune def. syndr. hum. retrovirol. 1998;18:270-276. 104. Lemp GF, Porco TC, Hirozawa AM, Lingo M, Woelffer G, Hsu LC, et al. Projected incidence of AIDS in San Francisco: The peak and decline of the epidemic. J. acquir. immune def. syndr. hum. retrovirol. 1997;16:182-189. 105. CDC. 1993 Revised Classification System for HIV Infection and Expanded Surveillance Case Definition for AIDS Among Adolescents and Adults. Morb. Mortal. Wkly. Rep. 1992;41:1-19. 106. Mulder DW, Nunn AJ, Kamali A, Naklylngi J, Wagner HU, Kengeya-Kayondo JF. Two-year HIV-1-associated mortality in a Ugandan rural population. Lancet 1994;343:1021-1023. 107. Wintrobe WM, Richard Lee G, Boggs DR, Bithell TC, Foerster J, Athens JW, et al. Clinical Hematology. 8th ed. Philadelphia: Lea & Febiger; 1981. 108. Monsuez JJ, Dufaux J, Vittecoq D, Flaud P, Vicaut E. Hemorheology in asymptomatic HIV-infected patients. Clin. Hemorheol. Microcirc. 2000;23:59-66. 109. Morfeldt-Manson L, Bottiger B, Nilsson B, von Stedingk LV. Clinical signs and laboratory markers in predicting progression to AIDS in HIV-1 infected patients. Scand. J. Infect. Dis. 1991;23:443-9. 110. Arango CA, Midani S, Alvarez A, Kubilis PS, Rathore MH. Usefulness of acute phase reactants in the diagnosis of acute infections in HIV-infected children. Southern Med. J. 1999;92:209-13. 111. Lefrere JJ, Salmon D, Doinel C, Rouger P, Courouce AM, Lambin P, et al. Sedimentation rate as a predictive marker in HIV infection. AIDS 1988;2:63-4. 112. Christie H. Interview with Dr. Robert Gallo July 1st Palexpo Conference Centre Geneva. [Betacam]. New York; 1998. 113. Kurth R, Teich NM, Weiss R, Oliver RTD. Natural human antibodies reactive with primate type-C antigens. Proc. Natl. Acad. Sci. U S A 1977;74:1237-1241. 114. Gallagher RE, Gallo RC. Type C RNA Tumor Virus Isolated from Cultured Human Acute Myelogenous Leukemia Cells. Science 1975;187:350-353. 115. Barbacid M, Bolognesi D, Aaronson SA. Humans have antibodies capable of recognizing oncoviral glycoproteins: Demonstration that these antibodies are formed in response to cellular modification of glycoproteins rather than as consequence of exposure to virus. Proc. Natl. Acad. Sci. U S A 1980;77:1617-1621. 116. Snyder HW, Fleissner E. Specificity of human antibodies to oncovirus glycoproteins: Recognition of antigen by natural antibodies directed against carbohydrate structures. Proc. Natl. Acad. Sci. U S A 1980;77:1622-1626. 117. Papadopulos-Eleopulos E. A Mitotic Theory. J. Theor. Biol. 1982;96:741-758. 118. Papadopulos-Eleopulos E. Reappraisal of AIDS: Is the oxidation caused by the risk factors the primary cause? Med. Hypotheses 1988;25:151-162. 119. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Kaposi's sarcoma and HIV. Med. Hypotheses 1992;39:22-9. 120. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM. Oxidative stress, HIV and AIDS. Res. Immunol. 1992;143:145-8. 121. Papadopulos-Eleopopulos E, Turner VF, Papadimitriou JM, Hedland-Thomas B, Causer D, Page B. A critical analysis of the HIV-T4-cell-AIDS hypothesis. Genetica 1995;95:5-24. 122. Papadopulos-Eleopopulos E, Turner VF, Papadimitriou JM, Causer D. Factor VIII, HIV and AIDS in haemophiliacs: an analysis of their relationship. Genetica 1995;95:25-50.
|